Questa macromolecola che forma i cromosomi e contiene l’informazione dell’essere vivente, ha la struttura di una doppia elica nella quale le spirali sono collegate l’una all’altra mediante coppie di basi nucleotidiche attraverso deboli legami a ponte idrogeno.
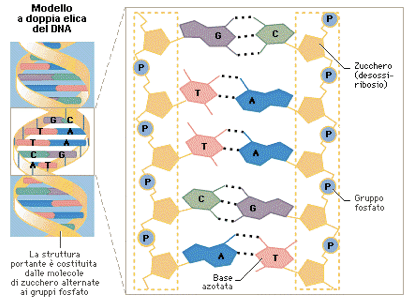
Questo filamento molecolare si trova raggomitolato in modo ordinato, riparato nel nucleo cellulare. È possibile, utilizzando bromuro di etidio, determinare lo srotolamento del DNA. Poiché il DNA funziona come un fotoaccumulatore, lo srotolamento determina una maggior emissione di biofotoni.
Dati sperimentali hanno confermato questa ipotesi.
Ci si aspetta che col procedere del grado evolutivo aumenti anche il contenuto del DNA. La quantità del DNA viene espressa in picogrammi (1pg=10-12g) e definita coefficiente C. Analizzando più di mille specie viventi, non si sono messe in evidenza significative differenze; si definisce questo inaspettato risultato: “paradosso del coefficiente C”.
Il DNA consta di una parte biologicamente molto attiva (eucromatina) utilizzata come matrice nella sintesi proteica e della restante parte ritenuta geneticamente inattiva (eterocromatina). La parte geneticamente attiva del DNA negli organismi superiori ammonta a circa il 2% del totale; pertanto le differenze tra la parte attiva e inattiva sono da attribuire essenzialmente all’eterocromatina (DNA inattivo). Il fatto che il 98% del DNA sia costituito da materiale di “scarto” risulta alquanto strano. Va dunque ricercata un’altra soluzione.
I risultati sperimentali di Popp, indicano che il DNA è la molecola centrale dalla quale procedono la regolazione biologica e l’emissione fotonica. Le basi nucleotidiche costituiscono efficaci fotoaccumulatori, adatti come ottimo materiale laser, infatti possiedono un ancoraggio stabile e un’elevata periodicità reticolare delle coppie di basi. Quando una delle basi assorbe luce, lo stato di attivazione che ne segue normalmente decade subito allo stato normale emettendo un fotone. Anche a temperatura ambiente, può accadere che invece di decadere allo stato normale, la molecola normale, si accoppi con una molecola vicina (generalmente attivata anch’essa) a formare una nuova unità: l’eccimero. Non appena l’eccimero, riemettendo un fotone, torna al suo stato primitivo, i due partner, che erano precedentemente collegati mediante il fotone, si respingono e decadono nei propri stati normali (monomeri), ad uno stato energetico più basso. Nei sistemi aperti l’inevitabile dispersione fotonica viene continuamente compensata mediante rinnovata attivazione degli eccimeri decaduti. Questo apporto di energia (pompaggio), deve compensare nell’aggregato cellulare l’energia dispersa della radiazione cellulare ultradebole. Lo stato di eccitazione dei monomeri viene raggiunto mediante pompaggio, per esempio di energia chimica proveniente dagli alimenti. La trasformazione in eccimeri soddisfa le condizioni laser.
La radiazione emessa dal DNA è in relazione con il numero degli eccimeri attivati. Il DNA agisce non solo come fonte ma anche come deposito di luce coerente (ordinata). Il risucchio di luce (nei calcoli definita potenziale fotochimico), permette di non attivare tutte le coppie di nucleotidi, ma di raggiungere un livello di soglia laser.
Erwin Schrodinger (premio Nobel per la fisica nel 1933), postulava un tale principio per i sistemi biologici già nel 1944, quando non era stato scoperto né il laser, né il DNA. Egli scrisse:”l’artificio mediante il quale un organismo si mantiene stabile ad un livello di ordine abbastanza elevato, consiste in realtà in un continuo assorbimento di ‘ordine’ dal suo ambiente”.
Dai calcoli risulta che la carica fotonica di una molecola di DNA è sufficiente per caricare circa 100.000 volte il potenziale della membrana cellulare. Di conseguenza sarebbe da considerare come il vero centro energetico del trasporto. L’attrazione tra le cellule adiacenti nell’aggregato cellulare potrebbe essere dovuta, secondo questo modello, al reciproco “risucchio” di fotoni che decide il segno e le forze attrattive.
L’elica del DNA si contrae quando riceve ed accumula fotoni in eccesso, poiché l’attivazione degli eccimeri induce sempre forze di attrazione tra le molecole. Scarsità di fotoni significa quindi espansione.
Tali contrazioni ed espansioni possono essere indotte dalla luce, la quale le eccita in senso oscillatorio.
Le molecole del DNA non sono soltanto puri elementi biochimici ma anche e soprattutto antenne capaci di emettere ed assorbire onde
Gli esperimenti dimostrano che di giorno, sotto l’azione della luce, il DNA si presenta effettivamente più denso che di notte. Mediante condensazione nella luce e decondensazione nell’oscurità, il DNA è in grado di formare intorno a sé un campo fotonico relativamente stabile, che a sua volta stabilizza il sistema stesso.
Impulsi caotici disturbano il DNA-laser ad eccimeri, interrompono il “contatto radio” con le cellule adiacenti, svuotano gli accumulatori fotonici, aumentando l’emissione fotonica.
È noto che la cellula si può trovare in 2 stadi fondamentali: l’interfase e la mitosi. L’interfase è l’arco di tempo compreso tra la fine della mitosi e l’inizio di essa; comprende a sua volta 3 sottofasi: G1, S e G2.
La fase G1, segue immediatamente la mitosi, è una fase di crescita cellulare durante la quale vengono elaborate proteine costitutive (es. della membrana cellulare), viene sintetizzato RNA messaggero, transfert e ribosomiale per garantire la sintesi proteica. In questa fase si osserva una intensificata emissione fotonica, contemporaneamente un forte assorbimento di calore e un’inibizione dell’accrescimento del tessuto circostante. È probabile che i decadimenti nella fase G1 siano dovuti a conversioni da singoletti a tripletti, possibili concause delle punte massime di intensità biofotonica.
Mentre gli eccimeri a singoletti garantiscono la coesione del DNA (cromosomi), gli eccimeri a tripletti probabilmente causano la parziale decondensazione del DNA (per favorirne la copiatura). I fotoni che in questa fase fuoriescono così intensamente probabilmente vanno ad attivare altre molecole del nucleo cellulare.
Nella seguente fase S, la cellula va incontro ad un’attività di sintesi con crescita e duplicazione del DNA (in preparazione alla mitosi in cui si devono generare 2 cellule figlie). Quando la cellula raggiunge un certo rapporto nucleo/citoplasma entra in uno stato di non ritorno. In questo momento c’è lo start, in cui la cellula è ormai costretta ad effettuare tutte le operazioni di mitosi. Applicando il modello biofotonico a questa fase si suppone si verifichi una “riattivazione fotonica nel proprio campo biofotonico”. Si suppone che i tripletti attivati stabilizzino le coppie di basi attraverso i ponti idrogeno. Quando questi eccimeri decadono nel loro stato normale, immediatamente si producono forze repulsive che spezzano i legami deboli a ponte idrogeno e impediscono un nuovo accoppiamento (filamento è aperto). La fessura creata risucchia le mobili ed agili basi nucleotidiche complementari presenti nel nucleo. Fotoni a polarizzazione circolare possono possono contribuire a dare il giusto impulso rotatorio per la despiralizzazione del DNA durante la duplicazione. I biofotoni in eccesso emessi dai filamenti del DNA costringono, per emissione indotta, i segmenti non ancora duplicati a passare nello stato di eccimero normale, finché al termine della fase S si è duplicata la molecola di DNA.
Nella fase G2 la cellula si prepara ad affrontare la fase mitotica e vengono duplicati gli organuli cellulari. Dal punto di vista biofotonico avviene il processo contrario rispetto alla G1: condensazione del Dna mediante trasformazione da tripletti a singoletti. La strutturazione produce più calore (entropia), risucchiando però contemporaneamente biofotoni. Il sistema supera di nuovo la soglia di laser. La potenza di pompaggio aumenta (glucosio, ATP, luce solare). L’energia dei fotoni solari di circa 550 nm è sufficiente ad attivare eccimeri a tripletti (350 nm) trasformandoli in monomeri a singoletti. Da questo stadio possono passare nello stato di eccimeri a singoletti. L’emissione di fotoni, che qui disturberebbe, si riduce; e si osserva un risucchio.
La struttura del campo assume in sequenza tutta la serie di distribuzioni conosciute di “modi” permessi, cominciando con gradi di attivazione elevati fino al livello più basso.
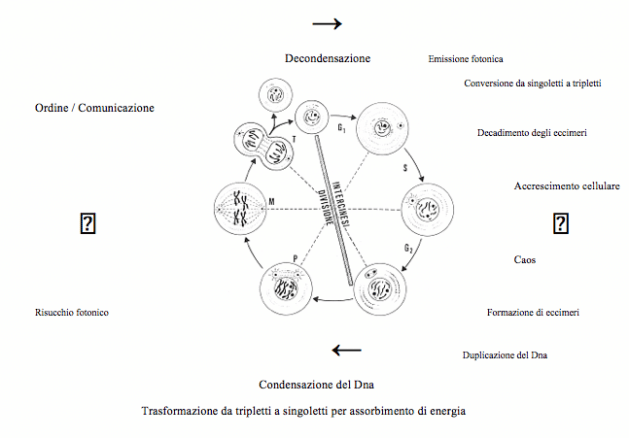
Mitosi cellulare e teoria dei biofotoni